 新闻
新闻科研进展
赖氨酸乙酰化修饰是重要的蛋白质翻译后修饰之一,通常指的是乙酰基团从乙酰辅酶A(Acetyl-COA)转移到蛋白质特定的赖氨酸 -氨基上,形成乙酰化的赖氨酸。赖氨酸乙酰化通常受到赖氨酸乙酰转移酶和去乙酰化酶的调控,从而改变蛋白的结构与功能,对细胞代谢、转录活性、蛋白质稳定性、信号通路等众多重要的生理功能进行精细的调节与控制。中国科学院水生生物研究所葛峰研究员和赵进东院士团队前期发现蓝细菌中很多蛋白都会发生乙酰化修饰,其中光合系统II蛋白PsbO的乙酰化修饰可以负调控光合放氧(Molecular & Cellular Proteomics, 2017, 16(7):1297-1311);蓝细菌中的乙酰化修饰受到去乙酰化酶CddA的调控,该酶能够通过去除底物蛋白的乙酰化修饰参与蓝细菌的光合作用过程(Plant Physiology, 2020, 184(2):762-776)。由于赖氨酸乙酰化修饰是可逆的,那么,蓝细菌中是否存在乙酰转移酶?
为回答这一重要科学问题,该团队对模式蓝细菌Synechococcus sp. PCC 7002中16个预测的乙酰转移酶进行系统筛选,通过体内和体外实验首次鉴定到一个乙酰转移酶cGNAT2,该基因敲除后导致蓝细菌的光合作用受到显著影响,提示cGNAT2可能通过调控底物蛋白乙酰化修饰参与蓝细菌的光合作用进程。
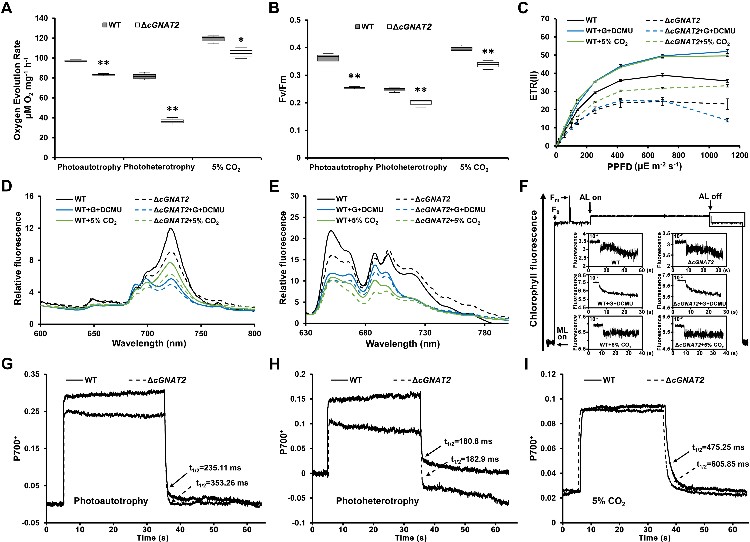
图1. 蓝细菌赖氨酸乙酰转移酶cGNAT2的功能研究
为了阐明cGNAT2潜在的催化机制,该团队利用AlphaFold2预测了cGNAT2的结构,发现该蛋白是一个同源二聚体的结构,由7个 折叠和4个 螺旋结构组成,形成一个管状口袋,能够与底物蛋白以及Acetyl-COA结合进而发挥催化作用;通过分子对接进一步确定了cGNAT2与Acetyl-COA之间的结合区域,并结合位点突变与体外酶活性实验证实了有8个氨基酸残基是维持酶活性的关键位点(图2)。
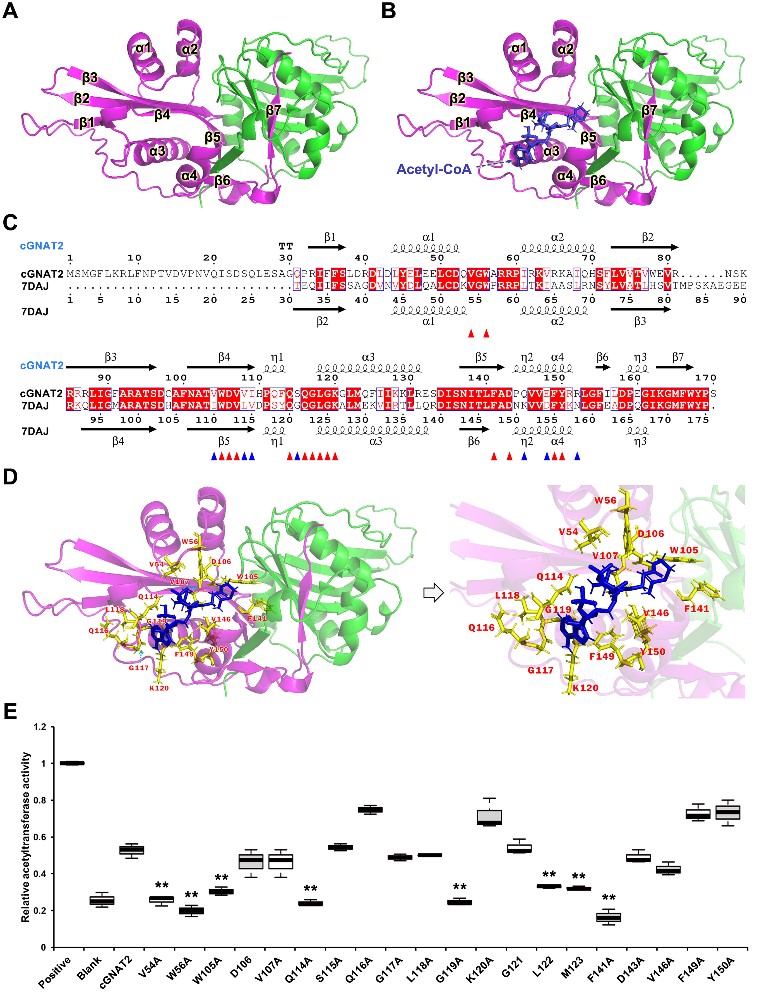
图2. cGNAT2的结构与催化活性
为了揭示cGNAT2的分子作用机制,通过采用乙酰化抗体富集和非标记定量乙酰组学技术,系统鉴定到548个cGNAT2调控的内源性靶标修饰蛋白,这些蛋白广泛分布于光合作用以及能量代谢通路中,表明cGNAT2调控的乙酰化修饰在光合系统以及能量代谢等生物学过程中发挥着重要的调控作用(图3)。
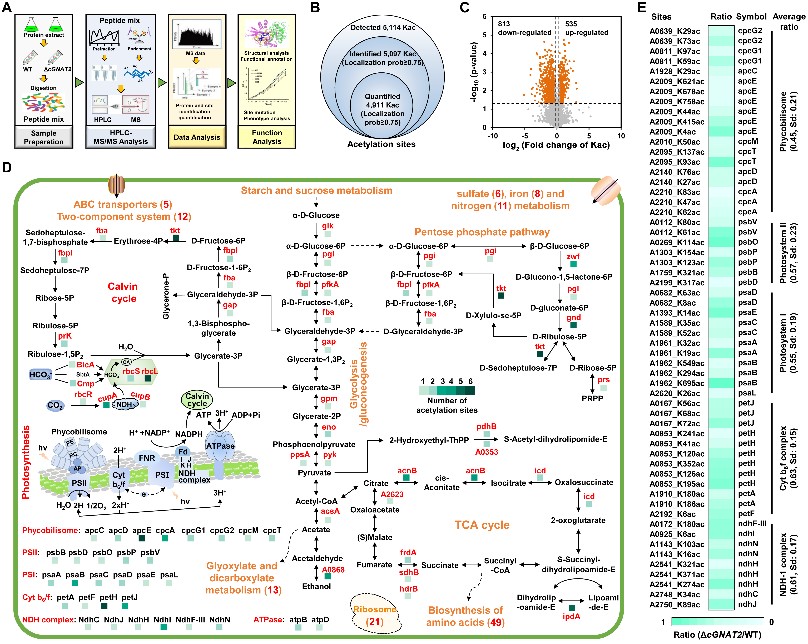
图3. cGNAT2调控的内源性靶标蛋白鉴定
进一步的功能实验发现,在光合作用通路中NdhJ(NDH脱氢酶亚基J)是cGNAT2的靶标底物,cGNAT2能够在体内和体外催化NdhJ的第89位乙酰化修饰,进而调控NdhJ的酶活性,并影响光合电子传递(图4)。
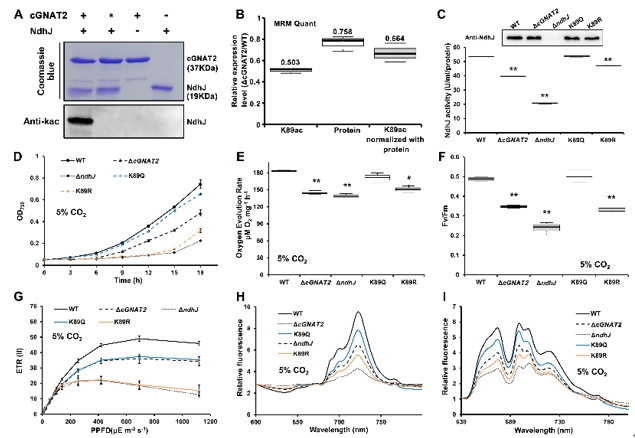
图4. cGNAT2调控NdhJ蛋白的乙酰化修饰进而影响光合作用过程
该研究首次在蓝细菌中发现了赖氨酸乙酰转移酶cGNAT2,并阐明了cGNAT2在蓝细菌中通过调控底物蛋白的乙酰化修饰进而实现其功能的分子机制(图5),对理解蛋白翻译后修饰在光合作用过程中的调控机理具有重要意义。
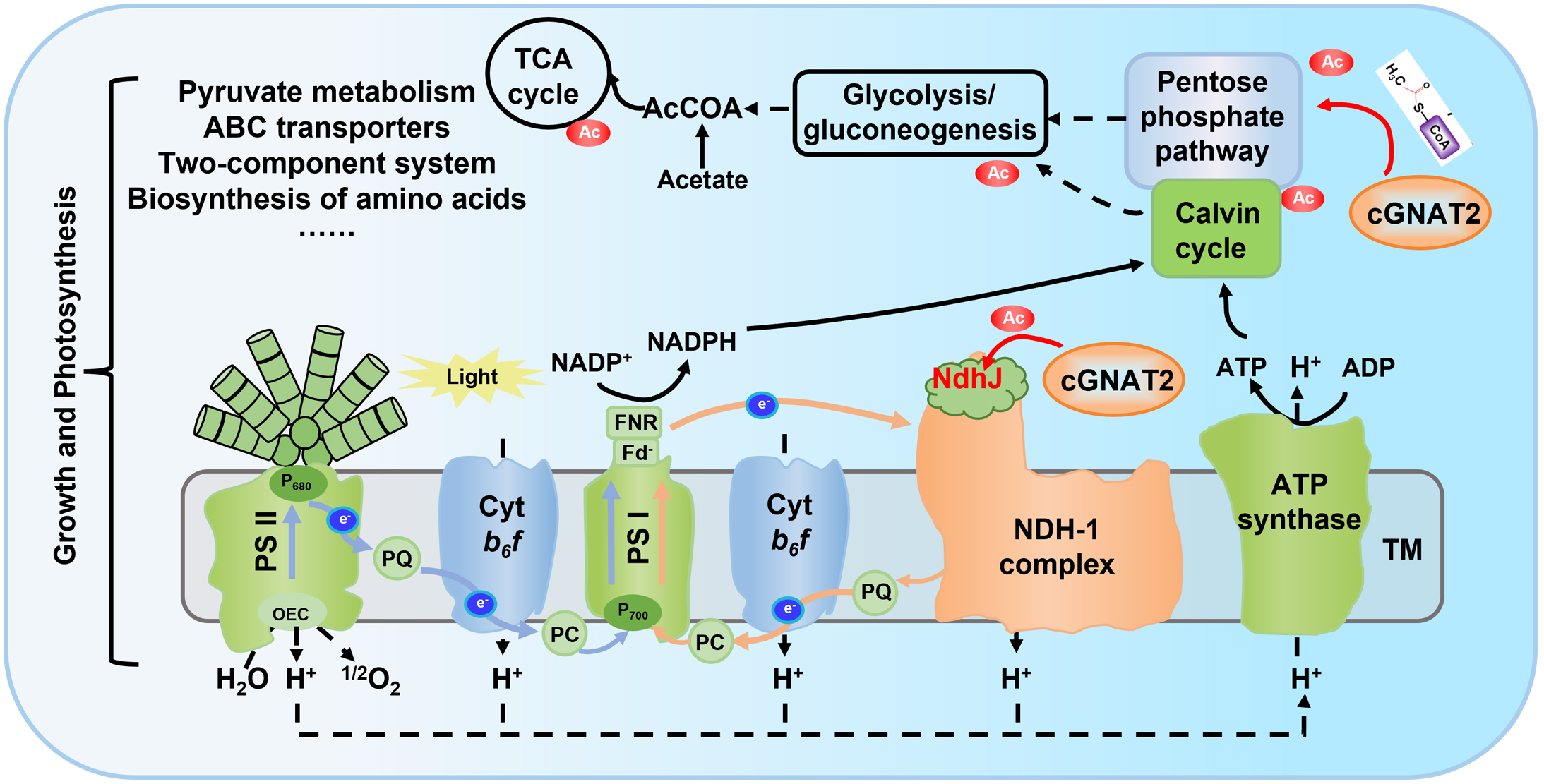
图5. cGNAT2在蓝细菌中调控生长和光合作用的分子机制模型
该研究成果“Deciphering structure, function, and mechanism of lysine acetyltransferase cGNAT2 in cyanobacteria”在线发表于Plant Physiology杂志,博士生贾坤、高级实验师杨明坤博士为该论文的并列第一作者,葛峰研究员、赵进东院士为共同通讯作者,该研究得到自然科学基金和国家重点研发计划的资助。文章链接:https://doi.org/10.1093/plphys/kiad509